

LINKED PAPER
Split moult: time constraints or endogenous strategy? Kiat, Y. & Izhaki, I. 2020. IBIS. DOI: 10.1111/ibi.12897 VIEW
‘Fine feathers make fine birds’. However, to ensure these fine feathers, birds need to replace them regularly. Old feathers become worn due to different environmental influences, potentially affecting the fitness of the birds (Jenni & Winkler 1994). That is why adult songbirds moult their complete plumage at least once a year. This process requires a lot of energy and usually occurs in the post-breeding period, just before the birds start migrating. In some species, however, the annual moult of flight feathers is divided over several periods or locations, such as summer and wintering areas. For example, the Common Whitethroat (Sylvia communis) moults some secondary feathers before autumn migration and renews other feathers during winter in a complementary moult. The mechanisms behind this pattern – known as ‘split moult’ – are a matter of debate among ornithologists (Nikolaus & Pearson 1991).
Schools of thought
There are two main schools of thought to explain the occurrence of split moult. On the one hand, Norman (1991) argued that interrupted moult of secondary feathers can be explained by time constraints. Species that breed at northern latitudes or that undertake long-distance migrations do not have the time to moult all their flight feathers and split their moult over two periods. Hedenström and his colleagues (1992), on the other hand, suggested that the interruption of moult is a strategy that is endogenously controlled by physiological processes within the bird. To discriminate between these two explanations, Yosef Kiat and Ido Izhaki compiled and analysed a database of moult patterns for more than 64,000 specimens, representing 213 species.
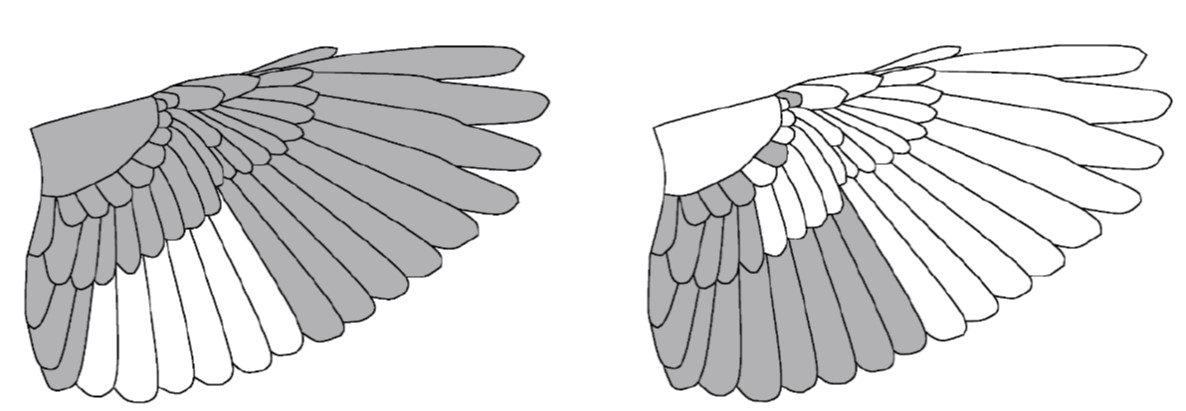
Figure 1 Example of a split moult strategy (moulted feathers in grey). The left drawing shows interrupted secondary feather moult during the main moult period. The right drawing shows complementary moult the non-moulted secondaries later in the annual cycle.
Experiments
The analyses provided evidence for interrupted moult in 36 passerines. Among long-distance migrants interrupted moult occurred in species that have their main moulting period in summer, following the breeding season. Interrupted moult was less common among species that moult in winter, before the breeding season starts. These patterns suggest that split moult is determined by time constraints because birds moulting after breeding are pressed with time before they start migrating. This conclusion is supported by an experiment on the Western Orphean Warbler (Sylvia hortensis) where researchers controlled the environmental conditions for 23 individual birds (Berthold & Querner 1982). Birds caught during the breeding season showed interrupted moult. But when the birds were prevented from breeding, they moulted several weeks earlier and completed their moult before migration. This suggests that the moulting pattern of the Western Orphean Warbler is not endogenously controlled, but driven by time constraints. Whether this explanation can be extrapolated to other species with interrupted moult remains to be tested with more experiments.
References
Berthold, P. & Querner, U. (1982). On the control of suspended moult in an European trans-Saharan migrant, the Orphean Warbler. Journal of the Yamashina Institute for Ornithology 14: 157–165. VIEW
Hedenström, A., Bensch, S., Hasselquist, D., Lindström, Å., Åkesson, S. & Pearson, D. (1992). Split moult: Stress or strategy? Ringing & Migration 13: 179–180. VIEW
Jenni, L. & Winkler, R. (1994). Moult and ageing of European passerines. A&C Black, London. VIEW
Nikolaus, G. & Pearson, D. (1991). The seasonal separation of primary and secondary moult in Palaearctic passerine migrants on the Sudan coast. Ringing & Migration 12: 46–47. VIEW
Norman, S.C. (1991). Suspended split‐moult systems‐an alternative explanation for some species of Palearctic migrants. Ringing & Migration 12: 135–138. VIEW
Image credits
Top right: Wing of Common Whitethroat Sylvia communis | Kiat & Izhaki (2020) Ibis
