


Gene flow and island endemism in the Buff-banded Rail
LINKED PAPER
Interisland gene flow among populations of the buff-banded rail (Aves: Rallidae) and its implications for insular endemism in Oceania. Garcia-R, J.C., Joseph, L., Adcock, G., Reid, J. & Trewick, S.A. 2017. Journal of Avian Biology. DOI: 10.1111/jav.01201. VIEW
The bird genus Gallirallus (Rallidae) includes widespread volant and local endemic flightless species on islands in the western Pacific (Fig. 1A). Although superficial similarity of insular species hints at their shared ancestry of flightlessness (e.g., Beauchamp, 1989) and parsimony-based ancestral state reconstruction of some traits supports this inference (Garcia-R et al., 2014), we argue that biology and landscape history render this unlikely. The islands occupied by flightless species are widely spaced and of varying age and geology (e.g., Neall and Trewick, 2008). Phylogenetic analysis of some of the many endemics and all of the volant species supports numerous independent origins of insular flightless taxa (Garcia-R et al., 2014, Garcia-R et al., 2016), several of which have been placed in different genera (e.g. Nesoclopeus, Habroptila, Diaphorapteryx) (Fig. 1B).
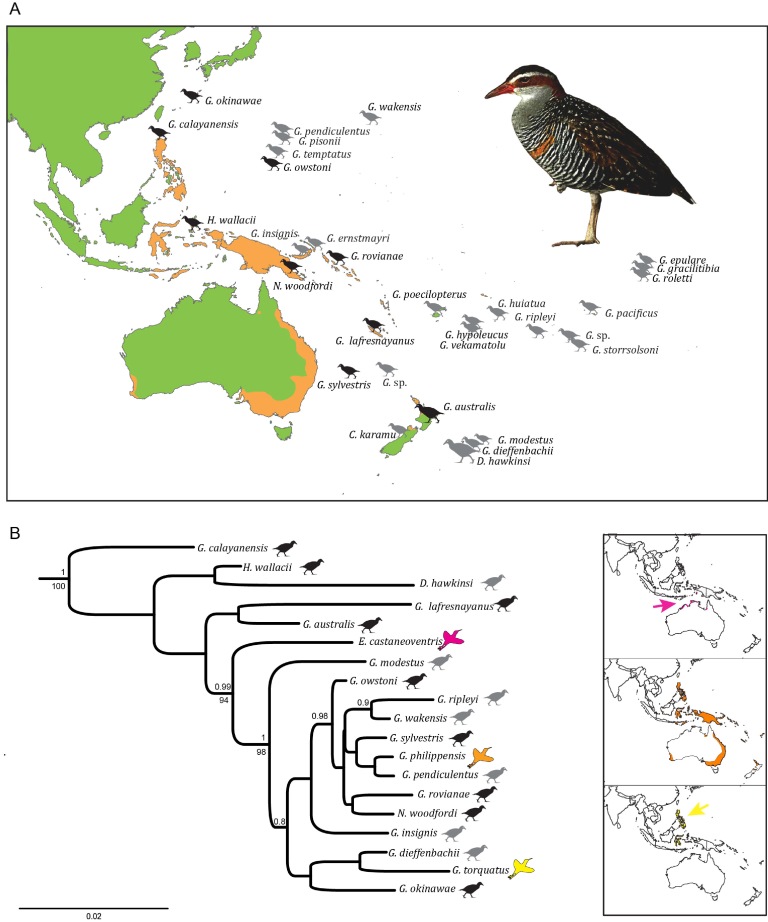 Figure 1 (A) Living (black) and extinct (gray) flightless insular species within Gallirallus in Western Pacific. Orange shading indicates the geographical distribution of the most widespread volant species Gallirallus philippensis. (B) Bayesian phylogenetic tree from concatenated alignment of DNA sequences reveals the importance of flightlessness in Gallirallus. Flighted species are shown with a winged-bird icon and flightless species with a walking-bird icon. Coloured winged-bird icons correspond to geographic range maps of volant species (right). Click on figure to view larger
Figure 1 (A) Living (black) and extinct (gray) flightless insular species within Gallirallus in Western Pacific. Orange shading indicates the geographical distribution of the most widespread volant species Gallirallus philippensis. (B) Bayesian phylogenetic tree from concatenated alignment of DNA sequences reveals the importance of flightlessness in Gallirallus. Flighted species are shown with a winged-bird icon and flightless species with a walking-bird icon. Coloured winged-bird icons correspond to geographic range maps of volant species (right). Click on figure to view larger
Diversification in Gallirallus primarily appears to be the result of repeated evolution of flightlessness on islands. As a result, flightless species look different from their flighted relatives but similar to each other (Diamond, 1991, Trewick, 1997b, Trewick, 2011). While it is obvious that flightlessness reduces the likelihood of further gene flow among island populations, this condition must first evolve in the face of potential gene flow among populations of the progenitor. A temporally uneven rate of gene flow and/or intense local selection would be needed for the observed outcome (Trewick, 1997a).
The most geographically widespread of the Gallirallus group is the volant Buff-banded Rail, Gallirallus philippensis (Fig. 2). It comprises over 20 subspecies distributed from the Philippines (~ 18o latitude North) in southeast Asia to subantarctic Macquarie Island in the Southern Ocean (~ 55o latitude South) in Oceania, and from the Cocos islands (~ 97o longitude East) in the Indian Ocean to Samoa (~ 172o longitude West) in the Pacific Ocean. The broad distribution of the Buff-banded Rail over large bodies of water shows a considerable propensity for dispersal and colonisation among islands, but its migratory habits are not well known. Subspecies of Buff-banded Rail differ mainly in plumage colours and are geographically separate in different archipelagos (Ripley, 1977, Taylor, 1998, Schodde and Naurois, 1982, Elliott, 1987), and this predicts spatial genetic structure caused by limited gene flow.
 Figure 2 Buff-banded Rail, Gallirallus philippensis | Aviceda | CC BY-SA 3.0
Figure 2 Buff-banded Rail, Gallirallus philippensis | Aviceda | CC BY-SA 3.0
In order to discern pathways of colonisation in the Western Pacific and patterns of gene flow among geographically structured groupings of the widespread Buff-banded Rail, we used population genetic analyses and coalescent models that help understand the role of selection in divergence and speciation within lineages. We found that genetic diversity tended to decrease and genetic differentiation (FST) increase as the geographic distance separating populations increased but haplotype diversity remained more even. However, both nucleotide and haplotype diversities were positively correlated with land area. Microsatellite data for a subset of locations showed moderate to high genetic differentiation and significant pairwise FST despite a relatively high migration rate (Fig. 3).
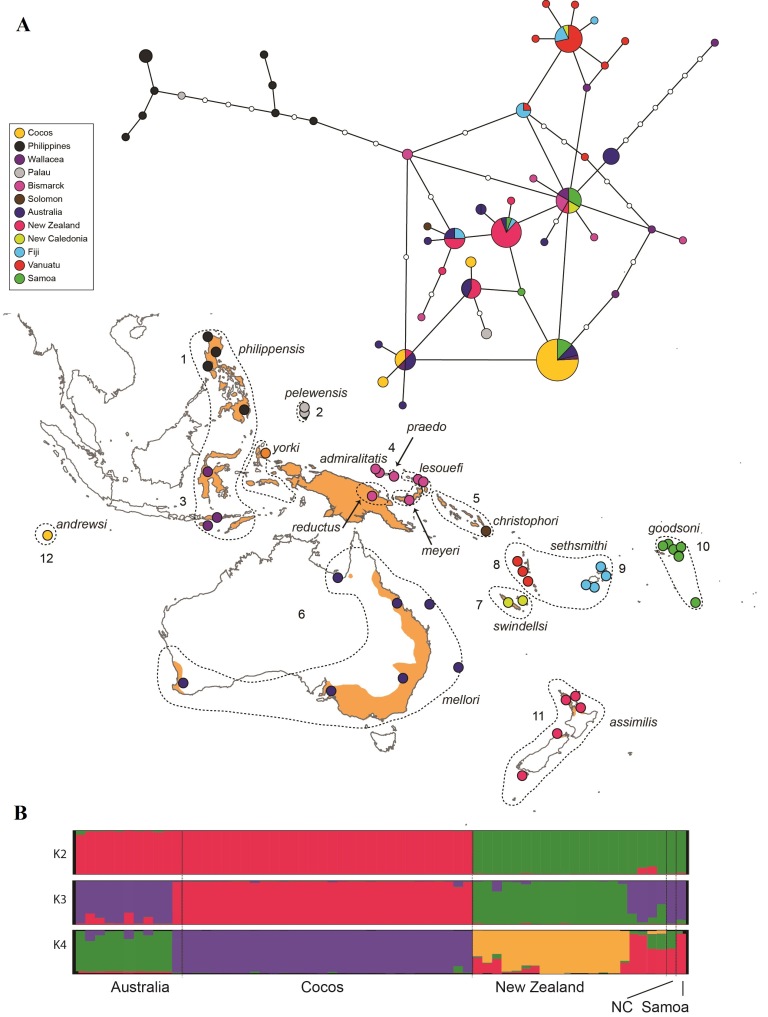 Figure 3 Distribution of genetic diversity in the volant Buff-banded Rail Gallirallus philippensis. (A) Minimum spanning network of mtDNA Control Region haplotypes. Colours correspond to 12 major geographical regions representing different archipelagos. (B) Genotypic variation among a subset of populations of Buff-banded Rail in Oceania surveyed using microsatellite loci. Click on figure to view larger
Figure 3 Distribution of genetic diversity in the volant Buff-banded Rail Gallirallus philippensis. (A) Minimum spanning network of mtDNA Control Region haplotypes. Colours correspond to 12 major geographical regions representing different archipelagos. (B) Genotypic variation among a subset of populations of Buff-banded Rail in Oceania surveyed using microsatellite loci. Click on figure to view larger
Our results are mostly consistent with a model of abrupt genetic changes due to founder events with multiple dispersals into Australia from Wallacea and the Bismarck Archipelago, and from Australia to other Pacific islands. Estimated high levels of gene flow, population sizes and similar divergence times among populations from coalescent analysis indicate that the key process in the formation of insular populations might be a strong selection pressure and not the loss of rare alleles by genetic drift during range expansion. The recognition of morphologically differentiated subspecies in the Buff-banded Rail (Schodde and Naurois 1982, Marchant and Higgins ) is an indication that divergent natural selection is not countered by the homogenizing effects of gene flow.
References
Beauchamp, A.J. 1989. Panbiogeography and rails of the genus Gallirallus. New Zealand Journal of Zoology 16: 763-772. VIEW
Diamond, J. 1991. A new species of rail from the Solomon islands and convergent evolution of insular flightlessness. The Auk 108: 461-470.
Elliott, G. 1987. Habitat use by the banded rail. New Zealand Journal of Ecology 10: 109-115. VIEW
Garcia-R, J.C., Elliott, G., Walker, K., Castro, I. & Trewick, S.A. 2016. Trans-equatorial range of a land bird lineage (Aves: Rallidae) from tropical forests to subantarctic grasslands. Journal of Avian Biology 47: 219-226. VIEW
Garcia-R, J.C., Gibb, G.C. & Trewick, S.A. 2014. Deep global evolutionary radiation in birds: Diversification and trait evolution in the cosmopolitan bird family Rallidae. Molecular Phylogenetics and Evolution 81: 96-108. VIEW
Neall, V.E. & Trewick, S.A. 2008. The age and origin of the Pacific islands: a geological overview. Philosophical Transactions of The Royal Society B 363: 3293-3308. VIEW
Ripley, S.D. 1977. Rails of the World: A Monograph of the Family Rallidae. David R. Godine Publisher, Boston, MA.
Schodde, R. & Naurois, R.D. 1982. Patterns of variation and dispersal in the Buff-breasted Rail (Gallirallus philippensis) in the South-west Pacific, with description of a new subspecies. Notornis 29: 131-141.
Taylor, B. 1998. Rails: A Guide to the Rails, Crakes, Gallinules and Coots of the World. Yale University Press, New Haven, Connecticut.
Trewick, S. 2011. Vicars and vagrants. Australasian Science 32: 22-25.
Trewick, S.A. 1997a. Flightlessness and phylogeny amongst endemic rails (Aves: Rallidae) of the New Zealand region. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 352: 429-446. VIEW
Trewick, S.A. 1997b. Sympatric flightless rails Gallirallus dieffenbachii and G. modestus on the Chatham Islands, New Zealand; morphometrics and alternative evolutionary scenarios. Journal of the Royal Society of New Zealand 27: 451-464. VIEW

Image credit
Featured image: Buff-banded Rail, Gallirallus philippensis | Toby Hudson | CC BY-SA 3.0
If you want to write about your research in #theBOUblog, then please see here.

