

LINKED PAPER
Multiple species within the Striated Prinia Prinia crinigera–Brown Prinia P. polychroa complex revealed by integrative taxonomy. Alström, P., Rasmussen, P.C., Sangster, G., Dalvi, S., Round, P.D., Zhang, R., Yao, C.-t., Irestedt, M., Le Manh, H., Lei, F. & Olsson, U. 2019. IBIS. DOI: 10.1111/ibi.12759. VIEW
Prinias, genus Prinia, comprise 23–26 species in the warbler family Cisticolidae (del Hoyo & Collar 2016, Gill & Donsker 2020). Two of the species, Red-winged Prinia P. erythroptera and Red-fronted Prinia P. rufifrons, were placed in the monotypic genera Heliolais and Urorhipis, respectively, until molecular phylogenetic analyses showed that they were nested within Prinia (Olsson et al. 2013). The same phylogeny showed that prinias are most closely related to tailorbirds Orthotomus spp., and proposed that these two genera should be placed in the subfamily Priniinae (Olsson et al. 2013).
The prinias are small to medium sized, with long, strongly graduated tails, discretely coloured plumages, mostly various shades of brown or grey, sometimes with some darker streaking, and frequently with pale and dark tail markings. Some of the species are rather similar in appearance. Their songs consist of monotonous fast repetitions of short phrases. The genus is widely distributed across Africa and southern Asia, and the species occur in dense vegetation such as undergrowth in open forest or along forest edges, bushes in open areas, or reedbeds. (Madge 2006).
Taxonomy of the Striated Prinia Prinia crinigera–Brown Prinia P. polychroa complex
The Striated Prinia Prinia crinigera–Brown Prinia P. polychroa complex is usually treated as two marginally sympatric species, the former found in southern and eastern Asia, and the latter in Southeast Asia, including Java (Madge 2006; Fig. 1; distributions modified based on our findings). The former occurs mainly in montane areas, whereas the latter is mostly found in the lowlands. Six or seven subspecies are usually recognised in P. crinigera and four in P. polychroa (Madge 2006, Gill & Donsker 2020).
Several authors have studied the taxonomy of various parts of this complex based on morphology alone (e.g. Sharpe 1903, La Touche 1922, Bangs 1930, Deignan 1942, Deignan 1957). However, there has been no consensus among authors regarding the number of species and subspecies or the circumscription of these. Some authors have treated all of the taxa as a single species (e.g. Vaurie 1959, Morony et al. 1975), whereas based on Deignan’s (1942, 1957) conclusions most recent authors have recognised two species (e.g. Madge 2006, Dickinson & Christidis 2014, del Hoyo & Collar 2016, Gill & Donsker 2020).
To revise the taxonomy of this complex, we undertook the first analysis integrating morphology, songs, mitochondrial and nuclear DNA, ecology and geographical distributions from throughout the range of the complex.
Based on morphology, we identified three main groups: (1) the crinigera group, comprising four subspecies from Pakistan to Yunnan Province, China; (2) the striata group, comprising three subspecies in China, and (3) the polychroa group, comprising four distinct populations, from Myanmar to central Vietnam and on Java, of which three subspecies were previously recognised (Fig. 1).
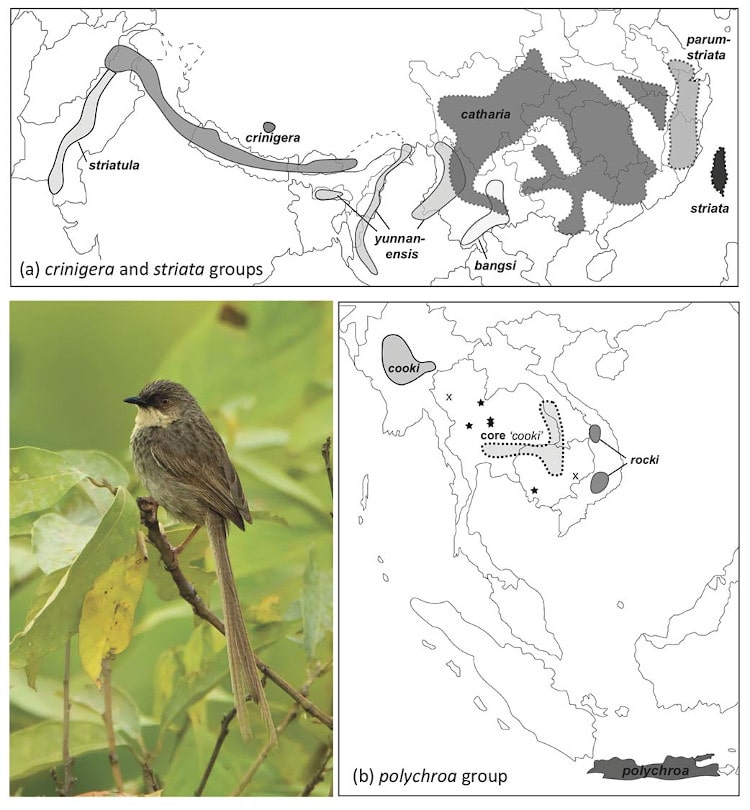 Figure 1 Distribution maps for taxa of the Prinia crinigera–P. polychroa complex: (a) the crinigera and striata groups, and (b) the polychroa group. Based on verified specimens, photographs and sound recordings, and using the taxon names that we recommend (except for ‘core cooki’, which we describe as a new taxon: P. polychroa deignani). In (a), taxa in the crinigera group have solid range borders, whereas these are dashed for taxa in the striata group. In (b), taxon ranges have solid borders except for ‘core cooki’, which is dashed for the main range. Stars indicate other ‘core cooki’ localities outside the main range. ‘X’ indicates records of uncertain taxon. Photo: P. crinigera yunnanensis, Pungro, Nagaland, May 2010 © Ramki Sreenivasan
Figure 1 Distribution maps for taxa of the Prinia crinigera–P. polychroa complex: (a) the crinigera and striata groups, and (b) the polychroa group. Based on verified specimens, photographs and sound recordings, and using the taxon names that we recommend (except for ‘core cooki’, which we describe as a new taxon: P. polychroa deignani). In (a), taxa in the crinigera group have solid range borders, whereas these are dashed for taxa in the striata group. In (b), taxon ranges have solid borders except for ‘core cooki’, which is dashed for the main range. Stars indicate other ‘core cooki’ localities outside the main range. ‘X’ indicates records of uncertain taxon. Photo: P. crinigera yunnanensis, Pungro, Nagaland, May 2010 © Ramki Sreenivasan
According to the phylogenetic analysis (Fig. 2), P. crinigera sensu lato (s.l.) comprised two non-sister groups of taxa (Himalayan crinigera and Chinese striata groups); the latter split from a common ancestor with the polychroa group c. 2.5 million years ago (mya). The polychroa group was divided into four clades representing rocki from Vietnam, cooki from Myanmar, birds from Cambodia and Thailand traditionally ascribed to cooki (here referred to as ‘core cooki’), and polychroa from Java. The oldest split within the polychroa group was between rocki and the others (c. 2 mya), whereas the youngest split, between ‘core cooki’ and polychroa was dated at 0.7 mya.
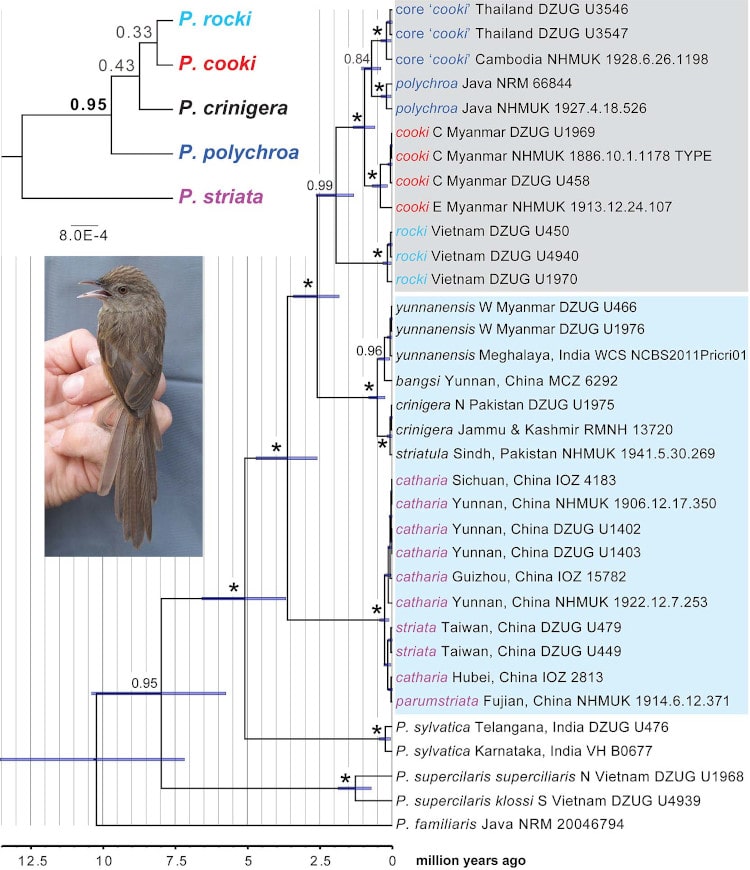 Figure 2 Mitochondrial cytochrome b (cytb) chronogram of the Prinia crinigera–P. polychroa complex, based on a molecular clock rate of 2.1%/million years, with 95% highest posterior density bars. The numbers at the nodes represent posterior probability (PP); *, PP = 1.00. Taxa traditionally treated as P. polychroa s.l. are shaded grey, and taxa traditionally treated as P. crinigera are shaded pale blue. Inset: *BEAST phylogeny based on cytb and nuclear ODC and myoglobin introns. The taxonomy in the inset phylogeny is the one we recommend based on the present study. Inset photo: Prinia polychroa ‘core cooki’ (described as a new taxon: P. polychroa deignani), presumed male, Sakaerat, Nakhon Ratchasima Prov., Thailand, 4 Jan. 2014 (DZUG U3546) © Philip D. Round
Figure 2 Mitochondrial cytochrome b (cytb) chronogram of the Prinia crinigera–P. polychroa complex, based on a molecular clock rate of 2.1%/million years, with 95% highest posterior density bars. The numbers at the nodes represent posterior probability (PP); *, PP = 1.00. Taxa traditionally treated as P. polychroa s.l. are shaded grey, and taxa traditionally treated as P. crinigera are shaded pale blue. Inset: *BEAST phylogeny based on cytb and nuclear ODC and myoglobin introns. The taxonomy in the inset phylogeny is the one we recommend based on the present study. Inset photo: Prinia polychroa ‘core cooki’ (described as a new taxon: P. polychroa deignani), presumed male, Sakaerat, Nakhon Ratchasima Prov., Thailand, 4 Jan. 2014 (DZUG U3546) © Philip D. Round
Two main types of songs were recorded: (1) long rattling series of repetitions of complex multi-note phrases (‘Type A’; Fig. 3) and (2) one or two pure notes repeated at longer intervals (‘Type B’; Fig 4). The crinigera and striata groups and Myanmar cooki produced only Type A songs. In contrast, ‘core cooki’ produced only Type B songs, whereas Vietnam rocki and Javan polychroa usually sang Type B songs, but rarely and briefly gave Type A song (both song types sometimes given by the same individual).
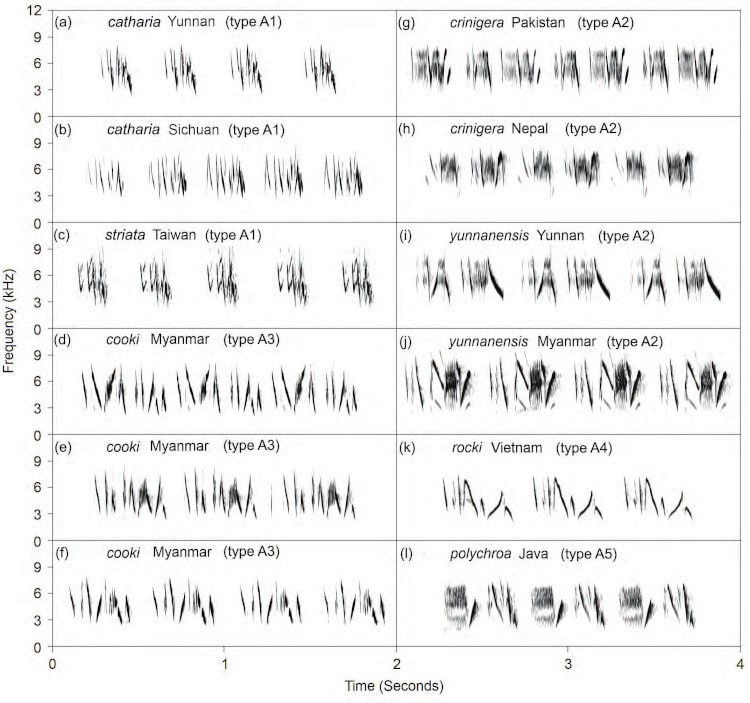 Figure 3 Sonograms of Type A songs of the Prinia crinigera–P. polychroa complex. This song type is rarely sung by taxa in the polychroa group (only noted in rocki and Javan polychroa). (a) catharia, Dali, Yunnan, 14 Jun. 2008 (AV19906); (b) catharia, Leibo, Sichuan, 30 May 2014 (AV19907); (c) striata, Taiwan (AV19919; Cheng-te Yao); (d) Myanmar cooki, Bagan, Myanmar, 12 Apr. 2000 (AV19908); (e) Myanmar cooki, Bagan, Myanmar, 12 Apr. 2000 (AV19909); (f) Myanmar cooki, Mt Popa, Myanmar, 22 Apr. 2000 (AV19910); (g) crinigera, Kaghan Valley, Pakistan, 7 Jun. 1998 (AV19902); (h) crinigera, Pulchowki, Kathmandu Valley, Nepal (Raf Drijvers, from Scharringa 2005); (i) yunnanensis, Liuku, Yunnan, 28 May 2008 (AV19903); (j) yunnanensis, Mt Victoria, Myanmar, 16 Apr. 2000 (AV19904); (k) rocki, Dalat, Vietnam, 12 May 1999 (AV19911); (l) Javan polychroa, Type A2, G. Halimun, W Java, Indonesia, 29 Oct. 2011 (AV19916; Bas van Balen). Recordings are by Per Alström unless otherwise noted.
Figure 3 Sonograms of Type A songs of the Prinia crinigera–P. polychroa complex. This song type is rarely sung by taxa in the polychroa group (only noted in rocki and Javan polychroa). (a) catharia, Dali, Yunnan, 14 Jun. 2008 (AV19906); (b) catharia, Leibo, Sichuan, 30 May 2014 (AV19907); (c) striata, Taiwan (AV19919; Cheng-te Yao); (d) Myanmar cooki, Bagan, Myanmar, 12 Apr. 2000 (AV19908); (e) Myanmar cooki, Bagan, Myanmar, 12 Apr. 2000 (AV19909); (f) Myanmar cooki, Mt Popa, Myanmar, 22 Apr. 2000 (AV19910); (g) crinigera, Kaghan Valley, Pakistan, 7 Jun. 1998 (AV19902); (h) crinigera, Pulchowki, Kathmandu Valley, Nepal (Raf Drijvers, from Scharringa 2005); (i) yunnanensis, Liuku, Yunnan, 28 May 2008 (AV19903); (j) yunnanensis, Mt Victoria, Myanmar, 16 Apr. 2000 (AV19904); (k) rocki, Dalat, Vietnam, 12 May 1999 (AV19911); (l) Javan polychroa, Type A2, G. Halimun, W Java, Indonesia, 29 Oct. 2011 (AV19916; Bas van Balen). Recordings are by Per Alström unless otherwise noted.
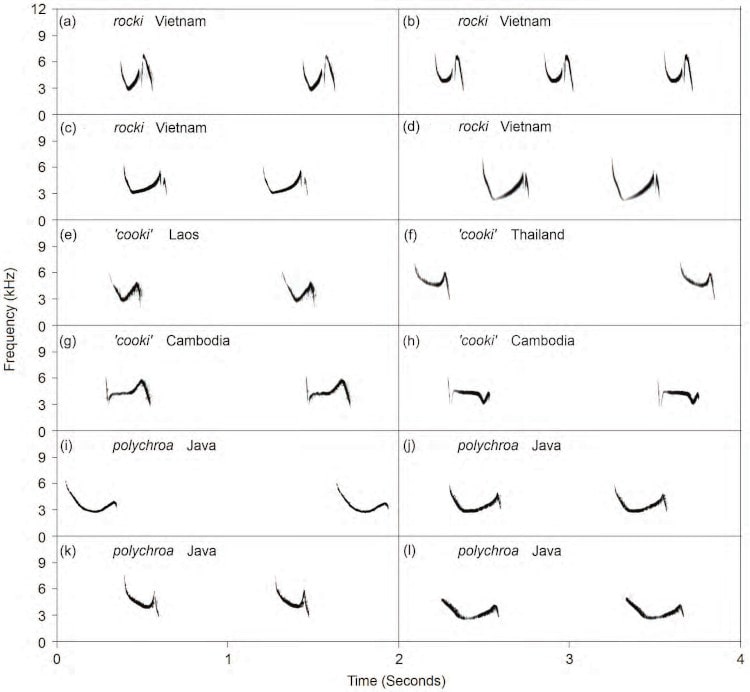 Figure 4 Sonograms of Type B songs of the P. polychroa complex. This is the usual song type of the polychroa group except Myanmar cooki, but it is not sung by any taxon of the crinigera or striata groups. (a) rocki, Dalat, Vietnam, 14 May 1999 (AV19912); (b) rocki, Dalat, Vietnam, 12 May 1999 (AV19914); (c) rocki, Dalat, Vietnam, 12 May 1999 (AV19913); (d) rocki, Dalat, Vietnam, 12 May 1999 (AV19915); (e) ‘core cooki’, Dong Khan Thung, Champasak Prov., south Laos (AV19920; Philip D. Round); (f) ‘core cooki’, Sabsadao Station, Thablan NP, Thailand, 3 Jul. 2014 (XC185665; Greg Irving); (g) ‘core cooki’, Tmat Boey, Preah Vihear Prov., Cambodia, Feb. 2008 (XC27624; David Edwards); (h) ‘core cooki’, Prey Veng, Preah Vihear Prov., Cambodia, 26 Dec. 2012 (XC124245; Patrik Åberg); (i) Javan polychroa, W Java, Indonesia, 14 Mar. 1986 (AV19905; Bas van Balen); (j) Javan polychroa, G. Halimun, W Java, 31 Oct. 2011 (AV19917; Bas van Balen); (k) Javan polychroa, G. Halimun, 29 Oct. 2011 (AV19916; Bas van Balen); (l) Javan polychroa, G. Halimun, 5 Nov. 2012 (XC142046; Frank Lambert). Note the two-note phrases in rocki. Recordings are by Per Alström unless otherwise noted.
Figure 4 Sonograms of Type B songs of the P. polychroa complex. This is the usual song type of the polychroa group except Myanmar cooki, but it is not sung by any taxon of the crinigera or striata groups. (a) rocki, Dalat, Vietnam, 14 May 1999 (AV19912); (b) rocki, Dalat, Vietnam, 12 May 1999 (AV19914); (c) rocki, Dalat, Vietnam, 12 May 1999 (AV19913); (d) rocki, Dalat, Vietnam, 12 May 1999 (AV19915); (e) ‘core cooki’, Dong Khan Thung, Champasak Prov., south Laos (AV19920; Philip D. Round); (f) ‘core cooki’, Sabsadao Station, Thablan NP, Thailand, 3 Jul. 2014 (XC185665; Greg Irving); (g) ‘core cooki’, Tmat Boey, Preah Vihear Prov., Cambodia, Feb. 2008 (XC27624; David Edwards); (h) ‘core cooki’, Prey Veng, Preah Vihear Prov., Cambodia, 26 Dec. 2012 (XC124245; Patrik Åberg); (i) Javan polychroa, W Java, Indonesia, 14 Mar. 1986 (AV19905; Bas van Balen); (j) Javan polychroa, G. Halimun, W Java, 31 Oct. 2011 (AV19917; Bas van Balen); (k) Javan polychroa, G. Halimun, 29 Oct. 2011 (AV19916; Bas van Balen); (l) Javan polychroa, G. Halimun, 5 Nov. 2012 (XC142046; Frank Lambert). Note the two-note phrases in rocki. Recordings are by Per Alström unless otherwise noted.
The Type A songs differed markedly among the crinigera and striata groups and Myanmar cooki. The Type B song differed clearly between rocki and the others, whereas ‘core cooki’ and polychroa were more similar to each other.
In conclusion, based on all available evidence, six main groups were identifiable: (1) the crinigera group (comprising the taxa striatula, crinigera, yunnanensis and bangsi); (2) the striata group (comprising catharia, parumstriata and striata); (3) Myanmar cooki; (4) ‘core cooki’; (5) rocki; and (6) Javan polychroa (cf. Figs 1 and 2). The crinigera and striata groups differed substantially morphologically, vocally and genetically (not even sisters), and were found to be broadly sympatric in Yunnan Province, China (yunnanensis and bangsi from the crinigera group and catharia from the striata group; Fig. 1). Myanmar cooki and rocki were each morphologically, vocally and genetically distinct. Javan polychroa and Thai, Cambodian and Laotian ‘core cooki’ differed slightly from each other in genetics, morphology and song.
Taxonomic implications
Based on our results, we recommend the recognition of five species in the complex, with the following suggestions for new English names: Himalayan Prinia P. crinigera (s.s.; with subspecies P. c. striatula, P. c. crinigera, P. c. yunnanensis and P. c. bangsi); Swinhoe’s Prinia P. striata (with subspecies P. s. catharia, P. s. parumstriata and P. s. striata; note that the name Chinese Prinia was suggested in the original paper, but that name was suggested for the recently split Prinia sonitans by del Hoyo and Collar 2016); Burmese Prinia P. cooki (monotypic); Annam Prinia P. rocki (monotypic); and Deignan’s Prinia P. polychroa (s.s.; with subspecies P. p. polychroa and the new P. p. deignani; see below).
As no name was available for the Thai, Cambodian and Laotian ‘core cooki’, it was described as P. p. deignani. It could be argued that because this taxon represents an evolutionarily distinct lineage, which is unlikely to come into secondary contact with the widely disjunct Javan polychroa, and which also differs from Javan polychroa in habitat choice (deignani is restricted to dry dipterocarp woodland, whereas Javan polychroa occurs in highly disturbed habitats), deignani could just as well be recognised as a distinct species. However, considering the recent divergence (0.7 mya) and slight morphological and vocal differentiation, we prefer to treat these as conspecific.
The taxon bangsi of Yunnan is part of the crinigera group rather than of the polychroa group, contra earlier statements (e.g. Deignan 1957), and hence there is no evidence for sympatry between P. polychroa s.l. and P. crinigera s.l., nor of the occurrence of P. polychroa in mainland China or Taiwan.
Acceptance of this revision implies the existence of three new single-country endemics: P. cooki from Myanmar, P. rocki from Vietnam and P. striata from China (mainland China and Taiwan).
This study highlights the importance of taxonomic revisions of poorly studied polytypic birds using a modern integrative taxonomic approach to better estimate the true diversity of bird species, and to understand their evolutionary history and biogeography.
References
Bangs, O. 1930. Types of birds now in the Museum of Comparative Zoology. Bull. Mus. Comp. Zool. 70: 147–426.
Deignan, H.G. 1942. A revision of the Indo-Chinese forms of the avian genus Prinia. Smiths. Misc. Coll. 103: 1–12.
Deignan, H. 1957. The races of the Longtail, Prinia polychroa (Temminck), with the description of a new race from southern Annam. Bull. Brit. Orn. Club 77: 24–25.
Dickinson, E.C. & Christidis, L. (eds) 2014. The Howard & Moore Complete Checklist of the Birds of the World: Fourth Edition, Vol. 2: Passerines. Eastbourne: Aves Press.
Gill, F. & Donsker, D. (eds) 2020. IOC World Bird List (v. 9.2). Available at: https://doi.org/10.14344/ioc.ml.9.2. Accessed on 1 January 2020. VIEW
del Hoyo, J. & Collar, N.J. 2016. HBW and BirdLife International Illustrated Checklist of the Birds of the World, Vol. 2: Passerines. Barcelona: Lynx Edicions.
La Touche, J.D. 1922. Mr. J. D. La Touche described the following new birds from S.E. Yunnan in S.W. China:— [Description of Suya crinigera parvirostris.]. Bull. Brit. Orn. Club 42: 51–55.
Madge, S.M. 2006. Prinia species accounts. In del Hoyo, J., Elliott, A. & Christie, D.A. (eds) Handbook of the Birds of the World, Vol. 11: 599–609. Barcelona: Lynx Edicions.
Morony, J.J., Jr, Bock, W.J. & Farrand, J. 1975. Reference List of the Birds of the World. New York: American Museum of Natural History.
Olsson, U., Irestedt, M., Sangster, G., Ericson, P.G.P. & Alström, P. 2013. Systematic revision of the avian family Cisticolidae based on a multi-locus phylogeny of all genera. Mol. Phylogenet. Evol. 66: 790–799. VIEW
Sharpe, R.B. 1903. A Hand-List of the Genera and Species of Birds, Vol. 4. London: British Museum.
Vaurie, C. 1959. The Birds of the Palearctic Fauna, Vol. 2. London: Witherby.

Image credit
Featured image: Prinia polychroa deignani, presumed male, Sakaerat Environmental Research Station, Nakhon Ratchasima Prov., Thailand, 4 Jan. 2014 © Philip D. Round

